联系人:苏先生
手 机:18612706349
电 话:010-57297418
传 真:010-57297418
邮 箱:suylinchcn@163.com
地 址:北京市北京经济技术开
发区荣华中路8号院13
号楼10层1111
邮 编:100176
[Handy PEA] 天然植物诱导剂2-氨基-3-甲基己酸保护茶树的生理活性免受高温损害

众所周知,高温(HT)是影响植物生理过程的最重要的限制因素之一。近年来随着全球气候变暖,极端天气发生次数逐渐增加,对植物生长和农业生产造成不可逆损害。中国茶叶栽培历史悠久,但夏季极端高温发生频率的增加已成为影响我国茶叶生产的重要限制因素之一。

2-氨基-3-甲基己酸(AMHA)是一种独特的天然植物抗高温胁迫诱导剂。南京农业大学生科院陈世国团队以温敏型茶树品种“龙井43”为材料,利用英国Hansatech公司生产的Handy PEA植物效率分析仪等仪器,研究了AMHA提高茶树夏季高温抗性的田间药效及其生理机制。研究结果“Natural plant inducer 2-Amino-3-Methylhexanoic acid protects physiological activity against high-temperature damage to tea (Camellia sinensis)”发表在Scientia Horticulturae(IF=4.342)杂志上。
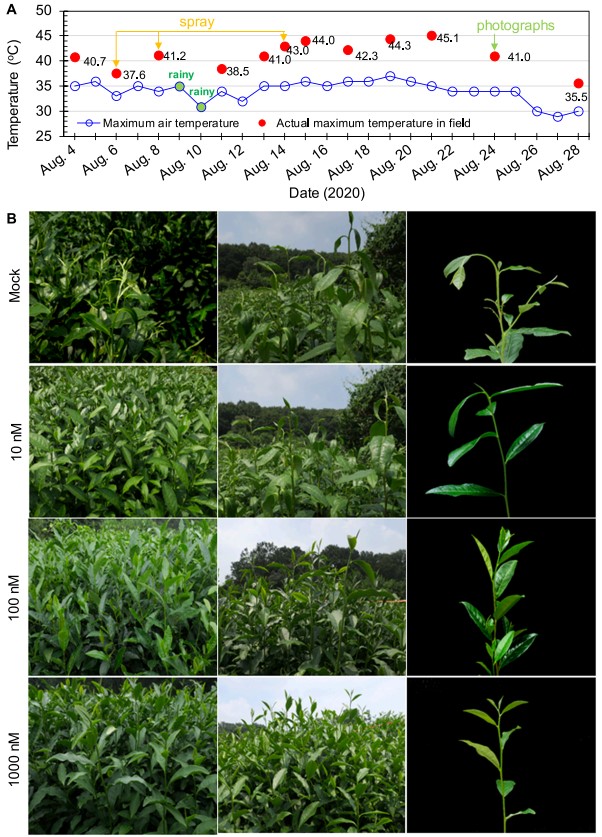
图1 (A)AMHA对大田茶树表型的影响。(B)在第三次施用不同浓度(10、100和1000nM)的AMHA模拟处理(0.02%Tween-20)10天后,田间茶树的表型照片。
由图1A可以看出,试验期间田间平均最高气温和实际最高气温分别在34℃和40℃以上。在第三次喷雾处理后7天,田间实际最高温度达到45.1℃(图1A)。在持续暴露于40℃以上的温度,特别是在8月21日遭受45.1℃的极端高温后,对照处理实验田中的许多茶树表现出严重的头部新枝下垂,甚至新芽和新叶萎蔫(图1B)。然而,在10nM AMHA处理的实验田的一些茶树中仅观察到新叶的轻微下垂。用100nM和1000nM AMHA处理的茶树具有直立的茎和正常的叶(图1B)。显然,外源施用极低剂量AMHA可以显著增强大田茶树对夏季高温的抗性。
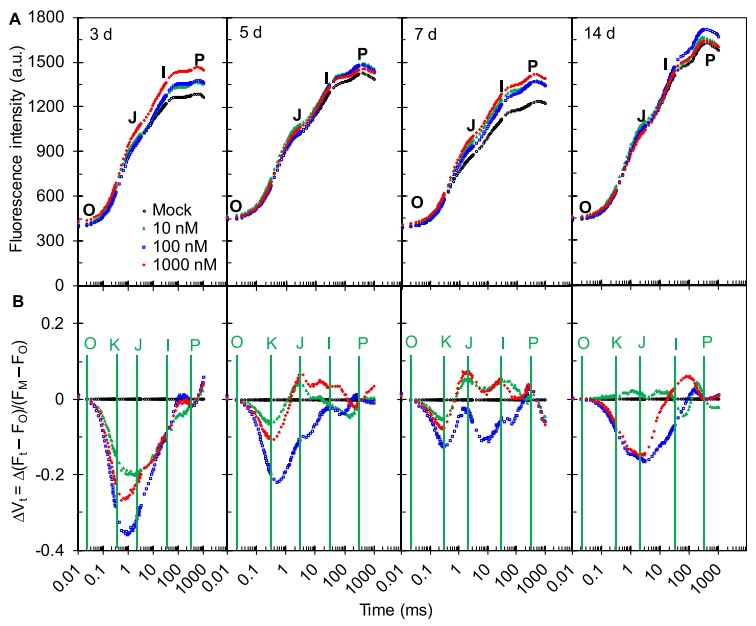
图2大田茶树施用AMHA后叶片OJIP曲线及振幅变化
为了研究AMHA在高温胁迫下提高光合速率的机制,在用不同浓度的诱导物处理后3d、5d、7d、14d测量茶树的荧光上升OJIP曲线(图2A)。AHMA处理的茶树的每条OJIP曲线的荧光产额均高于对照处理的茶树。可以看出,外源AMHA的施用缓解了高温对茶树荧光上升动力学的负面影响。为了获得比原始荧光动力学更详细的信息,将OJIP曲线进行双重归一化,ΔVt = Vt(处理)- Vt(对照)(图2B)。AMHA和对照处理之间ΔVt曲线的主要差异是出现负K峰。在100 nM AMHA下,与其他处理相比,还观察到较低的J峰(图2B)。有趣的是,100 nM AMHA比最高浓度的1000 nM AMHA对HT具有更好的保护作用(图2B)。
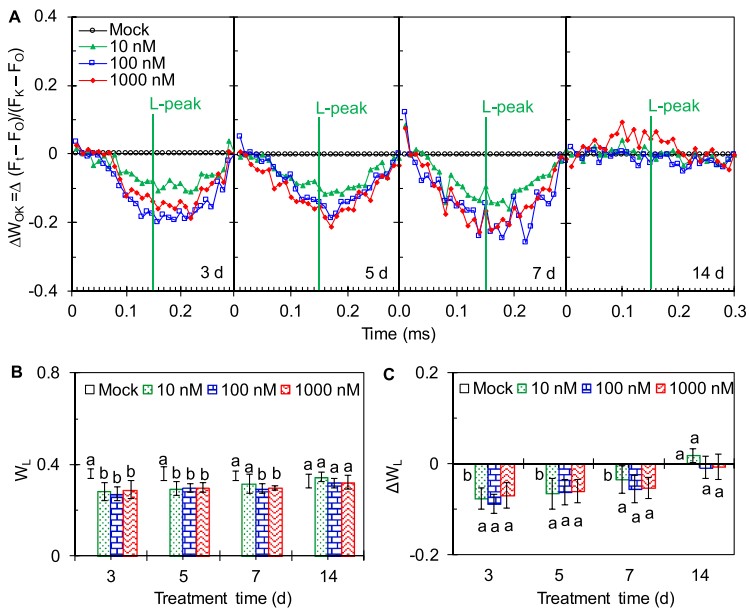
图3(A)AMHA施用对大田茶树OJIP曲线L阶的影响(A)线性时间标尺中的差异动力学ΔWOK = WOK(处理)− WOK(对照)表示为在OJIP曲线通过FO和FK双重归一化后显示L峰,WOK =(Ft-FO)/(FK-FO)。(B)L阶跃处的相对变量荧光与振幅FJ-FO的比值为WL =(FL-FO)/(FJ-FO)。(C)WL的差异为ΔWL =WL(处理)- WL(对照)。
为了进一步量化O-K阶段,将OJIP曲线在FO和FK进行双重标准化,使L峰可视化(图3A)。与对照相比,AMHA在3d、5d、7d时明显降低了L峰(图3A)。然而,其对L峰的影响在14d时消失(图3A),这意味着AMHA对PSII的保护作用消失。L峰变化的幅度使用WL=(FL-FO)/(FJ-FO)(图3B)和差值ΔWL=WL(处理)- WL(对照)(图3C)进行进一步定量。AMHA处理的茶树在3d、5d、7d时的WL和ΔWL值显著低于对照。WL和ΔWL值在3d时下降幅度最大,说明外源AMHA可以提高PSII内各结构之间的连接性,从而提高大田茶树对高温的抗性。
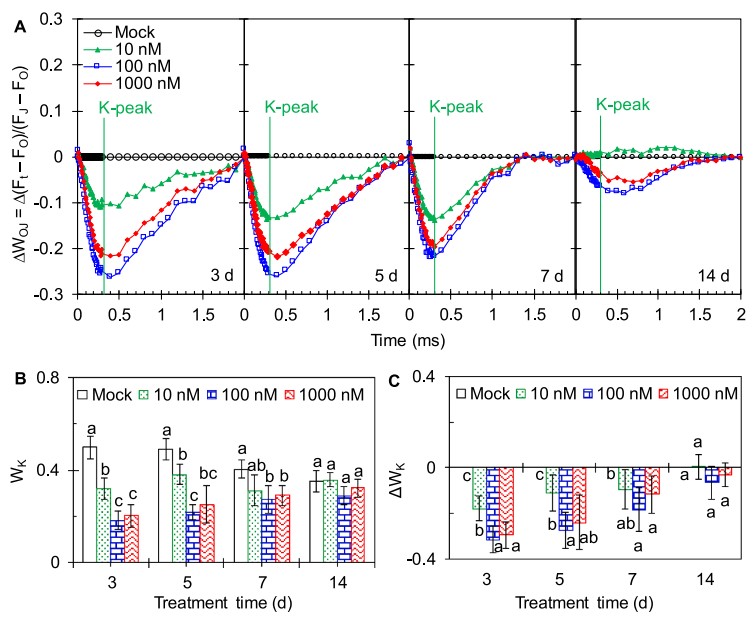
图4 AMHA对大田茶树OJIP曲线K阶跃的影响(A)线性时间标尺中的差异动力学Δ WOJ = WOJ(处理)− WOJ(对照)表示为在OJIP曲线通过FO和FJ双重归一化后显示K峰,WOJ =(Ft-FO)/(FJ-FO)。(B)K阶跃处相对变量荧光与振幅FJ-FO的比值,WK =(FK-FO)/(FJ-FO)。(C)WK差异为Δ WK = WK(处理)-WK(对照)。
图4A是对茶树OJIP曲线在O-J(20 μs-2 ms)之间进行的双重归一化结果。在AMHA处理中,茶树均出现了明显的ΔWOJ负K峰。AMHA对K峰的影响从第3天到第14天逐渐减弱。参数WK表示K阶的相对可变荧光(图4 B),ΔWK反映AMHA处理和对照茶树之间的WK差异(ΔWK =WK(处理)- WK(对照))(图4C),用于量化K峰。AMHA处理的茶树在3d、5d、7d时的WK和ΔWK值均显著低于对照,且随处理时间的延长而逐渐降低。WL和ΔWL值的最大降幅也出现在3d时,WK和ΔWK值的显著下降表明AMHA在施药后的前7 d能有效缓解高温对PSII放氧复合体OEC的破坏。
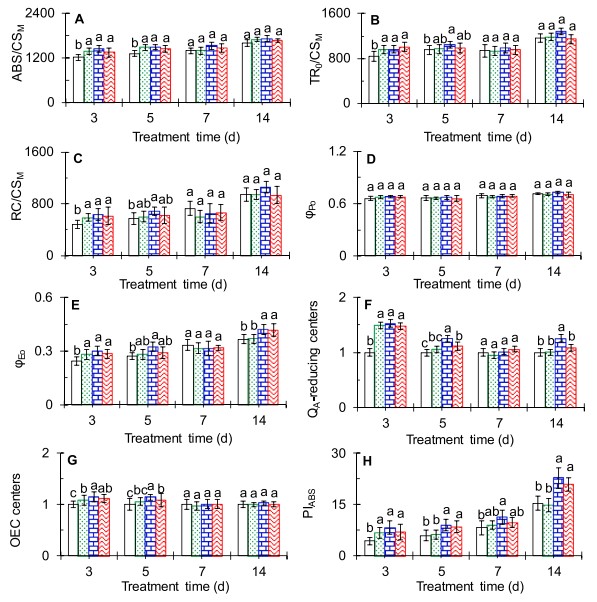
图5 AMHA施用对大田茶树选定的JIP test参数的影响AMHA处理中,每个激发叶片横截面的现象学能量通量ABS/CSM和TR0/CSM相对于模拟值显示出显著增加(图5A-5B)。此外,在整个试验期间,100 nM AMHA对ABS/CSM和TR0/CSM具有最积极的影响。同样,AMHA也导致RC/CSM显著增加(图5C)。从图5D中的数据可以看出,AMHA处理和模拟处理的茶树之间的φPo无显著差异。然而,AMHA能显著增加高温下茶树PSII电子传递的量子产率(φEo)(图5 E)。如图5F所示,AMHA导致QA-降低中心的值显著升高。AMHA增加了活性OEC中心的比例(图5G)。在3d时,与对照相比,分别增加了9%(10 nM)、15%(100 nM)和11%(1000 nM)。考虑到负K峰的结果(图4)并结合图5的荧光参数比较,可以看出外源AMHA的施用明显减轻了大田茶树活性OEC中心的高温伤害。
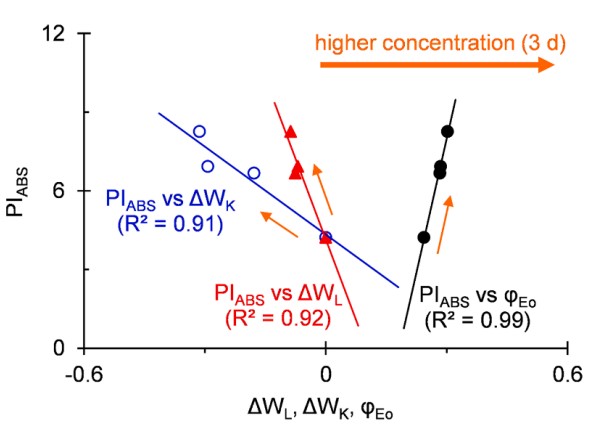
图6 PIABS与ΔWL、ΔWK和φEo的线性相关性分析
为阐明AMHA处理提高大田茶树光合作用的机理,分析了PIABS与其他JIP测试参数的关系。如图6所示,在不同浓度AMHA处理的茶树中,在3d时,PIABS与ΔWL或ΔWK之间观察到高度负相关,PIABS与φEo之间观察到显著正相关。这表明高温下AMHA处理PSII整体活性的上调主要是由于PSII和OEC状态的能量连接性改善以及PSII电子传递效率提高。显然,AMHA通过提高光合能力帮助大田茶树抵抗高温胁迫。
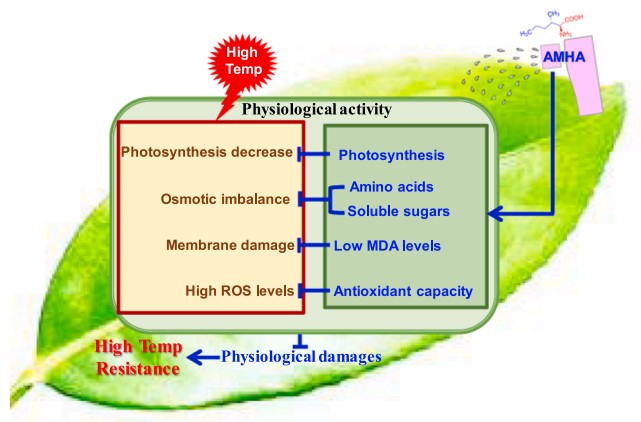
图7 AMHA诱导茶树对HT抗性的生理机制模型。
研究人员将本试验的发现整合为AMHA诱导茶树对高温抗性的生理机制模型(图7),并作出如下总结:高温破坏光合机构,产生过量的活性氧,引起氧化膜损伤,导致渗透压失衡,降低光合作用。外源AMHA主要通过提高茶树光合能力、增强抗氧化酶(POD、CAT和SOD)活性降低MDA含量、增加总氨基酸和可溶性糖含量从而维持渗透平衡、改善光合作用等途径缓解高温胁迫对茶树的生理伤害,最终提高茶树对高温胁迫的抗性水平。这些发现将有助于AMHA作为新型植物诱抗剂的开发与应用。
.png)

.jpg "全球同步发售,万物互联")




